No significant changes were observed in the surface endogenous GluR1 in the Tmub1/HOPS-RNAi neurons, as compared to the control neurons, suggesting that Tmub1/HOPS selectively regulates GluR2 and not GluR1. Similarly, interference with the NEEP21-GRIP or PICK1-GRIP interactions did not impair the surface expression of endogenous GluR1. Our results from the Tmub1/ HOPS overexpression experiments were consistent with those of the Tmub1/HOPS-RNAi experiments. The surface expression of GluR2, but not of GluR1, was increased in the Tmub1/HOPS-overexpressing neurons. Similarly, the expression of full-length GRIP enhanced the surface expression of coexpressed GluR2, suggesting that GRIP actively promotes GluR2 surface trafficking. Taken together, our immunostaining results and previous reports of the AMPAR surface staining suggest that the maintenance of the synaptic surface expression of GluR2 requires various interactions among non-PDZ proteins, PDZ proteins, and GluR2. Further, those interactions appear to affect the GluR2 subunit selectively. The amount of internalized GluR2 during 10 min did not differ significantly between the Tmub1/HOPS-RNAi and control neurons, suggesting that Tmub1/HOPS is not related to the endocytosis of GluR2 in the steady state. Recycling of the internalized GluR2 to the cell surface was significantly delayed in the Tmub1/HOPS-RNAi neurons, indicating that Tmub1/HOPS is related to the pathway by which GluR2 is recycled to the cell surface. Likewise, the inhibition of the NEEP21-GRIP and GRIP-PICK1 interactions delays the recycling of GluR2, suggesting that they are required for the recycling of GluR2 back to the plasma membrane. Consistent with our results of immunostaining of the surface endogenous GluR1, the internalization and recycling of GluR1 did not differ significantly between the Tmub1/HOPSRNAi and control 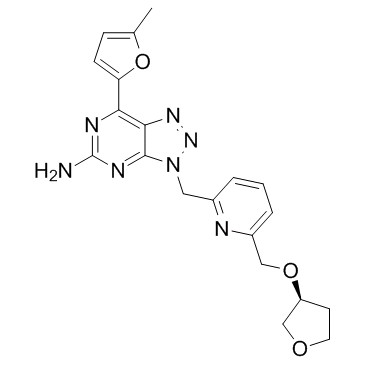 neurons. The inhibition of the NEEP21-GRIP interaction also did not affect the internalization and the recycling of GluR1. Therefore, Tmub1/HOPS appears to regulate the recycling of GluR2, which also requires multiple interactions such as those of NEEP21-GRIP and GRIP-PICK1. The results from the neurons overexpressing Tmub1/HOPS were consistent with those from the Tmub1/HOPS-RNAitransfected neurons. The internalization of GluR2 did not change but the recycling of GluR2 varied significantly, supporting that Tmub1/HOPS is related to the recycling of GluR2 but not to the internalization of GluR2. The level of GluR2 or GluR1 recycled to the surface in the control cells appeared to be relatively low as compared to that in other reports, although the time scales used by us were slightly different. In the absence of neuronal activity, internalized AMPARs are sorted for Homatropine Bromide either Mepiroxol degradation or reinsertion at synapses ; however, upon incubation with AMPA or NMDA/TTX, AMPARs are more actively directed into the recycling pathway. Because we did not use any artificial stimulation in our recycling assay, some AMPARs may spontaneously be sorted for lysosomal degradation, which was represented as a rather lower recycling ratio in the control cells, as compared to other reports. In surface-receptor regulation, Tmub1/HOPS appears to act contrary to ubiquitin and the UBL protein SUMO. While ubiquitin and SUMO decrease the surface number of their target receptors, Tmub1/HOPS increases the surface number of AMPARs. Like Tmub1/HOPS, some other UBL domaincontaining proteins, such as Plic-1/ubiquilin-1 and GABARAP/ ubiquilin-2, also increase the surface number of their receptors although the underlying mechanisms may be somewhat different.
neurons. The inhibition of the NEEP21-GRIP interaction also did not affect the internalization and the recycling of GluR1. Therefore, Tmub1/HOPS appears to regulate the recycling of GluR2, which also requires multiple interactions such as those of NEEP21-GRIP and GRIP-PICK1. The results from the neurons overexpressing Tmub1/HOPS were consistent with those from the Tmub1/HOPS-RNAitransfected neurons. The internalization of GluR2 did not change but the recycling of GluR2 varied significantly, supporting that Tmub1/HOPS is related to the recycling of GluR2 but not to the internalization of GluR2. The level of GluR2 or GluR1 recycled to the surface in the control cells appeared to be relatively low as compared to that in other reports, although the time scales used by us were slightly different. In the absence of neuronal activity, internalized AMPARs are sorted for Homatropine Bromide either Mepiroxol degradation or reinsertion at synapses ; however, upon incubation with AMPA or NMDA/TTX, AMPARs are more actively directed into the recycling pathway. Because we did not use any artificial stimulation in our recycling assay, some AMPARs may spontaneously be sorted for lysosomal degradation, which was represented as a rather lower recycling ratio in the control cells, as compared to other reports. In surface-receptor regulation, Tmub1/HOPS appears to act contrary to ubiquitin and the UBL protein SUMO. While ubiquitin and SUMO decrease the surface number of their target receptors, Tmub1/HOPS increases the surface number of AMPARs. Like Tmub1/HOPS, some other UBL domaincontaining proteins, such as Plic-1/ubiquilin-1 and GABARAP/ ubiquilin-2, also increase the surface number of their receptors although the underlying mechanisms may be somewhat different.