These puncta are essentially abolished by replacing the regulatory C-terminal tyrosine residue of Src by phenylalanine. In addition, mutations disrupting the SH3 binding motif in wild type PTP1B reveal a secondary site of interaction. Our results show for the first time physical interactions of ER-bound PTP1B with Src at puncta localized at the plasma Ginsenoside-F4 membrane in contact with the substrate, and further reveal that this interaction critically depends on the active site of PTP1B and the regulatory tyrosine 529 of Src. The results of the present paper illustrate a case of functional modulation in trans, among molecules located at the surface of the ER and plasma membrane, a phenomenon which may apply to a wide range of molecules and have impact in the regulation of several cell processes. In the present paper we used the BiFC technique to demonstrate direct physical interactions among ER-bound PTP1B and tyrosine kinases Src and Fyn at the plasma membrane. These interactions were revealed as bright fluorescence puncta associated with the ER. Our results strongly 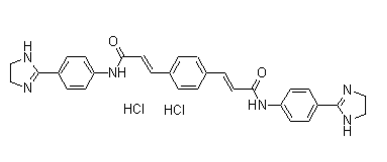 suggest that PTP1B, which is localized at the tip of dynamic ER tubules, was positioned close to the ventral membrane in contact with the substrate and interacted with Src at multiple puncta sites. We further show that mutations altering the active site of PTP1B and removing the negative regulatory residue of Src significantly reduced BiFC complex formation. These results suggest that ER-bound PTP1B releases Src from its negative regulation at random point contacts of the membrane/ substrate interface, leading to its activation and possibly recruitment to adhesion complexes. The subcellular localization of BiFC puncta is expected to be conditioned by the subcellular localization of each interaction partner. PTP1B is anchored to the cytosolic face of the ER membrane through a hydrophobic C-tail, allowing its mobility through the vast surface of the ER, and its interaction with substrates in the cytosol. In addition, dynamic changes of shape of the ER, dependent on its association with microtubules, extend the range of PTP1B interactions to a larger spatial scale, positioning PTP1B in the cell cortex, and therefore facilitating its encounter with substrates associated with the cytosolic face of the plasma membrane. Among these potential substrates, several laboratories including ours have identified the Src family of tyrosine kinases. Src kinases associate with plasma membrane by means of fatty acid modifications, and by protein protein interactions. In addition, a fraction of Src remains in the cytosol and another is associated with recycling endosomes. On this regard, the weak BiFC signal revealing the ER network, when using the wild type PTP1B, likely represents interactions with the freely diffusing pool of cytosolic Src. In contrast, bright BiFC puncta in the ER likely reflects interactions with spatially restricted Src and PTP1B molecules. We found that BiFC puncta did not co-localize with rab11, a recycling endosomes marker, and did not display directional movement in the cytoplasm, as expected for traffic carriers. Therefore, it is unlikely that BiFC puncta reflect interactions with an endosomal pool of Src. The fact that BiFC puncta were Cinoxacin retained in ventral membrane preparations, were visualized within the evanescent field produced by TIRF microscopy, and some colocalized with dark spots seen under SRIC microscopy, strongly suggest that they could represent spatially restricted interactions of ER-bound PTP1B with a subset of Src localized at the plasma membrane. Indeed, BiFC was significantly reduced when SrcTYN was used.
suggest that PTP1B, which is localized at the tip of dynamic ER tubules, was positioned close to the ventral membrane in contact with the substrate and interacted with Src at multiple puncta sites. We further show that mutations altering the active site of PTP1B and removing the negative regulatory residue of Src significantly reduced BiFC complex formation. These results suggest that ER-bound PTP1B releases Src from its negative regulation at random point contacts of the membrane/ substrate interface, leading to its activation and possibly recruitment to adhesion complexes. The subcellular localization of BiFC puncta is expected to be conditioned by the subcellular localization of each interaction partner. PTP1B is anchored to the cytosolic face of the ER membrane through a hydrophobic C-tail, allowing its mobility through the vast surface of the ER, and its interaction with substrates in the cytosol. In addition, dynamic changes of shape of the ER, dependent on its association with microtubules, extend the range of PTP1B interactions to a larger spatial scale, positioning PTP1B in the cell cortex, and therefore facilitating its encounter with substrates associated with the cytosolic face of the plasma membrane. Among these potential substrates, several laboratories including ours have identified the Src family of tyrosine kinases. Src kinases associate with plasma membrane by means of fatty acid modifications, and by protein protein interactions. In addition, a fraction of Src remains in the cytosol and another is associated with recycling endosomes. On this regard, the weak BiFC signal revealing the ER network, when using the wild type PTP1B, likely represents interactions with the freely diffusing pool of cytosolic Src. In contrast, bright BiFC puncta in the ER likely reflects interactions with spatially restricted Src and PTP1B molecules. We found that BiFC puncta did not co-localize with rab11, a recycling endosomes marker, and did not display directional movement in the cytoplasm, as expected for traffic carriers. Therefore, it is unlikely that BiFC puncta reflect interactions with an endosomal pool of Src. The fact that BiFC puncta were Cinoxacin retained in ventral membrane preparations, were visualized within the evanescent field produced by TIRF microscopy, and some colocalized with dark spots seen under SRIC microscopy, strongly suggest that they could represent spatially restricted interactions of ER-bound PTP1B with a subset of Src localized at the plasma membrane. Indeed, BiFC was significantly reduced when SrcTYN was used.