An engineered tau variant that mimics constitutively phosphorylated tau by replacing Ser/Thr residues with glutamate, did not develop tau aggregates, neuronal loss, or behavioral abnormalities. In contrast, mice expressing tauR406W, a variant that remains hypophosphorylated compared to tauwt even in aged mice, developed age-dependent tau aggregates and memory and behavioral abnormalities. The hypophosphorylated tauP301L species in neurospheres and young mice may represent free tau not associated with microtubules. Tau bound to MTs acquires more phosphorylation than free tau as demonstrated by in vitro phosphorylation of tau in the Folinic acid calcium salt pentahydrate presence or Pimozide absence of MTs. Many of the MAPT exon 10 missense mutations that cause dementia, including P301L, reduce the ability of tau to interact with MT, and tauwt displaces mutant tau from MTs. The absence of tauP301L aggregates or neurofibrillary tangles in neurospheres and in young mice, despite phosphorylation at many of the sites most frequently phosphorylated in AD and FTD, also may correspond to unbound tau as MT association has been implicated as an important step for tau nucleation. The quantity of NFTs correlates with disease severity. However, recent studies have dissociated NFTs from neuronal death and decreased memory function, and instead suggest a deleterious effect of soluble tau. Phosphorylation of mouse tau also reflects appropriate phosphorylation corresponding to the differentiation state. Heavily phosphorylated 3R mouse tau is know to bind to nucleolar organizing regions in dividing cells and was also observed in neurospheres, indicating that human tau did not inhibit either the cellular machinery or kinases involved. This reinforces the same conclusion coming from the recapitulation of the genetic differences in human tau phosphorylation seen in neurosphere culture. We observed differences in filopodia-spine densities between tauwt and tauP301L differentiated cells. Developmentally, dendritic spine morphology evolves from long thin filopodia-spines to mature spines of various morphologies; during this transformation, filopodia-spine density decreases. We observed a slightly higher filopodia-spine density in Map2 and TUJ-1 double-positive cells from tauP301L cultures than those derived from tauwt or cultures that did not express MAPT. While the level of transgene expression within individual cells did not affect filopodia-spine density, we have not ruled out an effect of the transgene insertion site. Interestingly, in a separate P301L mouse line that that harbors the mutation in the longest mouse tau isoform, 4R2N, driven by the Thy1 promoter, young mice with “hypophosphorylated” tau have enhanced learning and memory and increased Long Term Potentiation in the dentate gyrus 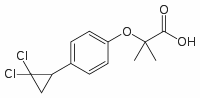 compared to controls. With age, the spine density of rTg4510 mice decreases and coincides with increased neuronal excitability. Whether or not the greater filopodiaspine density we observed in differentiated tauP301L cultures relates to enhanced LTP in young mice or neuronal vulnerability later in life in unknown, but warrants further exploration. We did not observe mislocalization of tauP301L to dendritic spines, as reported in aging mice or in transfected rat neuron cultures. We saw high levels of transgenic tau protein throughout neuritic projections in differentiated neurospheres from both human tau transgene genotypes. The presence of tau in the filopodia-spines of developing cells suggests that the phenomenon correlates with a dynamic morphology and does not necessarily indicate pathological injury.
compared to controls. With age, the spine density of rTg4510 mice decreases and coincides with increased neuronal excitability. Whether or not the greater filopodiaspine density we observed in differentiated tauP301L cultures relates to enhanced LTP in young mice or neuronal vulnerability later in life in unknown, but warrants further exploration. We did not observe mislocalization of tauP301L to dendritic spines, as reported in aging mice or in transfected rat neuron cultures. We saw high levels of transgenic tau protein throughout neuritic projections in differentiated neurospheres from both human tau transgene genotypes. The presence of tau in the filopodia-spines of developing cells suggests that the phenomenon correlates with a dynamic morphology and does not necessarily indicate pathological injury.