Mice lacking CCL3 have a reduced inflammatory response to influenza virus and are resistant to coxsackievirus-induced myocarditis. Although these chemokines can play positive roles in resolving acute inflammation, they are increasingly recognized as detrimental for chronic inflammation, and therapeutic inhibitors of MIP1inflammatory chemokines are being developed to treat diseases associated with chronic inflammation. The increased expression of these chemokines in kidneys infected with WT C. albicans, therefore, could play a role in the strong inflammatory responses we observed. In the absence of Dur1,2p activity, inflammation and expression of CCL3 and, more reproducibly, CCL4 was markedly decreased. Altered expression of MIP1 chemokines was recently associated with virulence in a different C. albicans mutant. Both chemokines showed decreased circulating levels in mice infected with C. albicans lacking HMX1, and their expression was regulated by the immunosuppressive carbon monoxide produced by Hmx1p. Decreased kidney expression of CCL3 and CCL4 was also noted in mice infected with a pmr1D mutant of C. albicans with decreased virulence. Determining how Dur1,2p expression increases CCL3 and CCL4 expression in infected kidneys, therefore, is an important topic for future research. Given the importance of the kidney for concentrating and excreting urea, we propose that urea metabolism to ammonia and CO2 by Dur1,2p plays a role in the persistence of C. albicans in this organ. CO2 is known to inhibit macrophage clearance of C. albicans, and so may limit the efficacy of the macrophages recruited into infected kidneys. NH3 has been considered as a virulence factor for C. albicans, and its importance for virulence is well established for Helicobacter pylori. NH3 produced by this bacterial pathogen via urease plays a critical role in controlling local pH and facilitating invasion through the gastric mucosa. A second enzymatic pathway that produces NH3 by deamidation of asparagine or glutamine is also essential for colonization of the stomach environment by H. pylori. NH3 plays an additional role in virulence of H. pylori by enhancing host cell apoptosis through its modulation of endocytic vesicle trafficking. Furthermore, NH3 is increasingly recognized as an important signaling molecule in cellular responses to LOUREIRIN-B stress. In the context of inflammation, elevated NH3 levels inhibit neutrophil chemotaxis, phagocytosis, and degranulation while also stimulating spontaneous oxidative bursts. NH3 also inhibited the capacity of neutrophils to engulf bacteria. Notably, the latter study implicated the p38 pathway in this neutrophil dysfunction, and we observed differential expression 4 MAP kinases that are direct or indirect activators of p38. Therefore, NH3 production by Dur1,2p may mediate some of the changes in host gene expression and neutrophil recruitment between mice infected with WT and the dur1,2 mutant. Our results are also compatible with the specific requirement of DUR1,2 for germ tube formation in the macrophage and evidence that that urea metabolism via Dur1,2p provides ammonia for external alkalinization of the C. albicans environment, thus permitting germ tube formation and escape from the phagolysosome. Despite its Albaspidin-AA defect in hyphal-dependent escape from macrophages, the dur1,2 mutant can make hyphae except when driven exclusively by arginine in macrophages or in response to urea in vitro. This selective defect in hyphal differentiation may contribute to the lower abundance 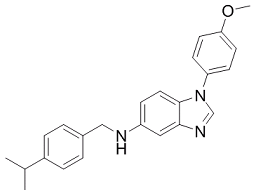 of filaments in kidneys infected with the mutant strain at 3 and 5 days PI. Taken together, our present and previous results strongly indicate that expression of Dur1,2p enhances kidney neutrophil infiltration but limits phagocytic clearance of C. albicans.
of filaments in kidneys infected with the mutant strain at 3 and 5 days PI. Taken together, our present and previous results strongly indicate that expression of Dur1,2p enhances kidney neutrophil infiltration but limits phagocytic clearance of C. albicans.
Month: June 2019
By EC and vascular SMC in response to pro-inflammatory stimuli in a NF-kB and activator protein dependent manner
Significantly heightened inflammatory responses detected in both EC and SMC at 12 and 24 H suggested a central role for inflammation in driving the Cinoxacin injury response to vein graft implantation. By 7 D, inflammation related genes tapered down without returning to basal levels. In fact, inflammatory genes spanned all time-points, were the most abundant of differentially regulated genes, and were likely the key regulators of other pathogenic processes. We propose that early targeting of key inflammatory molecules could be an ideal approach to prevent vein graft implantation injury. Previous in vivo studies have demonstrated that blocking inflammatory responses do attenuate IH. Following heightened inflammation driven injury response, we observed at later time-points, a significant upregulation of cell cycle related genes in both EC and SMC. Recovery of the endothelium through cell proliferation is beneficial to the healing process, while that of SMC may be deleterious, as it promotes the development of a neointimal layer, i.e. IH, the pathognomonic feature of mid-term vein graft failure. Accordingly, therapies aimed at preventing/treating vein graft failure using cell cycle inhibitors must spare EC, specifically target SMC, and ideally be effective for at least a week following vein graft implantation. Failure of the PREVENT trial may be related to the therapeutic agent not fulfilling all of these criteria. Genes involved in the extracellular matrix reorganization were significantly up-regulated by 30 D in both EC and SMC. In particular, collagen genes were the most enriched class of ECM components, consistent with the prominence of collagen as a constituent of IH lesion. Deposition of ECM is essential for vein graft healing in response to injury under normal circumstances, however in the special case of implantation injury it can lead to Mechlorethamine hydrochloride stenosis and graft failure. Delayed therapies that could tackle ECM deposition need to be optimized for reducing pathogenic vascular remodeling while promoting positive vascular remodeling. In order to delineate a causality relationship between differentially expressed genes after vein graft implantation, we analyzed the data using a backpropagation approach that integrates interactions between differentially expressed genes from the different points, starting at the latest time-point; i.e. 30 D. This means of analyzing the data offered a unique perspective of identifying the upstream pathogenic effectors of vein graft implantation injury, based on endpoint molecular signals involved in lesion formation. Furthermore, this approach allowed us to integrate into the same network genes derived from significantly affected biological pathways and define interconnectivity between these pathways. We identified 6 and 5 biological pathways that were dominant in the backpropagation networks of EC and SMC, respectively. Remarkably 4 of these pathways were common to both cell types, three of which spanned all time-points, namely the IL-8, IL-6 and dendritic cell maturation pathways. We surmise that these 3 pathways are not only critical pathogenic effectors of ![]() vein graft implantation injury, but also harbor promising therapeutic targets. Within these pathways, the IL-8 gene itself was up-regulated from 2 H to 7 D in EC and from 12 H to 7 D in SMC, and the IL-6 gene was up-regulated from 12 H to 7 D in both EC and SMC. We propose that both IL-8 and IL-6 are central to the pathogenesis of vein graft implantation injury. Specifically, IL-8 is a pro-inflammatory CXC chemokine produced mainly by neutrophils, monocytes and macrophages.
vein graft implantation injury, but also harbor promising therapeutic targets. Within these pathways, the IL-8 gene itself was up-regulated from 2 H to 7 D in EC and from 12 H to 7 D in SMC, and the IL-6 gene was up-regulated from 12 H to 7 D in both EC and SMC. We propose that both IL-8 and IL-6 are central to the pathogenesis of vein graft implantation injury. Specifically, IL-8 is a pro-inflammatory CXC chemokine produced mainly by neutrophils, monocytes and macrophages.
Interrogated the transcriptome in a temporal and cell type specific manner in order to better understand
In the present study, we used this canine model, the pathophysiology of vein graft implantation injury and identify novel therapeutic strategies. Specifically, we used Laser Capture Microdissection to isolate endothelial and medial layers for subsequent RNA extraction and transcription profiling. Our study focused on early transcriptional events that we believe to be key in dictating Folinic acid calcium salt pentahydrate outcome. We did not interrogate transcriptional changes of the neointima since this layer does not develop in this model, or in clinical vein grafts before 30 D. In fact IH is not significant in this model before 60�C90 days. Future work is focused on interrogating neointimal transcriptome in a longterm model of canine vein graft failure LCM yielded highly 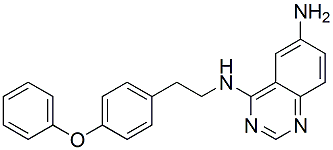 enriched EC and SMC samples allowing the study of cell specific transcriptional changes. To our knowledge, this is the first study using this approach. Unsupervised PCA analysis of our gene chips demonstrated that graft EC and SMC genes formed time dependent clusters with maximal transcriptional changes appearing 12 to 24 H following implantation, and diminishing over the 30 D time period. This tapered response suggests that early intervention during the peri-operative period may be sufficient to prevent/inhibit vein graft implantation injury, and therefore limit the development of IH. Supervised analysis of the transcriptional data depicted temporal modification of gene expression in EC and SMC. Interestingly, some genes were modified over continuous time-points, whereas others were either non-continuously modified or modified at a single time-point, highlighting the importance of defining optimal therapeutic windows for each given target. For example, the proinflammatory chemokine IL-8 was up-regulated in EC from 2 H up to 7 D, and in SMC from 12 H up to 7 D. This pattern of up-regulation in both cell types could be ideal for a targeted therapy aiming at interrupting early and mid pathogenic inflammatory culprits involved in vein graft implantation injury. In contrast, Col1A1 was up-regulated in both EC and SMC only at 30 D, suggesting that this extracellular matrix gene is likely a downstream participant in the pathogenesis of vein graft implantation injury, and key to the vascular remodeling that occurs in vein grafts. In addition this analysis also suggested that graft EC might be the first responders to vein graft implantation injury, as they obviate a change in their transcriptome as early as 2 H following implantation. In contrast, SMC do not show any significant changes at this early time-point. Gene ontology analysis on temporally, and differentially expressed genes indicated that implantation injury affected multiple ontological categories in both, EC and SMC. Enriched GO clusters included apoptosis, inflammatory and immune responses, mitosis and extracellular matrix reorganization. In EC, apoptosis related genes were up-regulated and peaked at 2 H following implantation confirming that rapid EC LOUREIRIN-B damage and loss was critical in triggering the cascade of events leading to vein graft injury. In contrast, apoptosis related genes were up-regulated and peaked in SMC at 12 H. The significance of increased expression of pro-apoptotic genes in medial SMC is still unclear. Several studies suggest that increased medial SMC apoptosis promotes vascular damage and influences the phenotypic switch of these cells from contractile to synthetic, therefore promoting IH.
enriched EC and SMC samples allowing the study of cell specific transcriptional changes. To our knowledge, this is the first study using this approach. Unsupervised PCA analysis of our gene chips demonstrated that graft EC and SMC genes formed time dependent clusters with maximal transcriptional changes appearing 12 to 24 H following implantation, and diminishing over the 30 D time period. This tapered response suggests that early intervention during the peri-operative period may be sufficient to prevent/inhibit vein graft implantation injury, and therefore limit the development of IH. Supervised analysis of the transcriptional data depicted temporal modification of gene expression in EC and SMC. Interestingly, some genes were modified over continuous time-points, whereas others were either non-continuously modified or modified at a single time-point, highlighting the importance of defining optimal therapeutic windows for each given target. For example, the proinflammatory chemokine IL-8 was up-regulated in EC from 2 H up to 7 D, and in SMC from 12 H up to 7 D. This pattern of up-regulation in both cell types could be ideal for a targeted therapy aiming at interrupting early and mid pathogenic inflammatory culprits involved in vein graft implantation injury. In contrast, Col1A1 was up-regulated in both EC and SMC only at 30 D, suggesting that this extracellular matrix gene is likely a downstream participant in the pathogenesis of vein graft implantation injury, and key to the vascular remodeling that occurs in vein grafts. In addition this analysis also suggested that graft EC might be the first responders to vein graft implantation injury, as they obviate a change in their transcriptome as early as 2 H following implantation. In contrast, SMC do not show any significant changes at this early time-point. Gene ontology analysis on temporally, and differentially expressed genes indicated that implantation injury affected multiple ontological categories in both, EC and SMC. Enriched GO clusters included apoptosis, inflammatory and immune responses, mitosis and extracellular matrix reorganization. In EC, apoptosis related genes were up-regulated and peaked at 2 H following implantation confirming that rapid EC LOUREIRIN-B damage and loss was critical in triggering the cascade of events leading to vein graft injury. In contrast, apoptosis related genes were up-regulated and peaked in SMC at 12 H. The significance of increased expression of pro-apoptotic genes in medial SMC is still unclear. Several studies suggest that increased medial SMC apoptosis promotes vascular damage and influences the phenotypic switch of these cells from contractile to synthetic, therefore promoting IH.
In the heterogeneous composition of transgene expressing and non-expressing cells within the same sphere
Unlike clonal cultures, these neurospheres were formed by aggregation, which results in culture heterogeneity.The minor transgene expression variability among Lomitapide Mesylate neurosphere cultures generated from littermate fetuses possibly occurred during the initial brain harvest. We did not specifically dissect the forebrain from each fetus. Nestin expressing neural SCs in the developing midbrain and hindbrain may have contributed to the transgene expressing cell population since CKIIa, the promoter driving tTA-transgenic human tau expression, is expressed throughout the brain at this developmental age. We saw more variability among independent experimental harvests than among cultures derived from littermate fetuses; we attribute this variation to inconsistencies inherent to IFA. Regardless, IFA consistently demonstrated that undifferentiated cells derived from rTg expressing fetal brains expressed human tau in a higher proportion of cells than those derived from rTg expressing fetal brains. Total brain homogenates Mechlorethamine hydrochloride indicate that rTg mice express comparable levels of transgenic tau as rTg. While the rTg cultures had a lower proportion of total cells expressing human tau, they contained a greater proportion of cells that expressed higher levels of tau. This feature of some transgenes is caused by position-effect variegation. Neurospheres, like the mice from which they were derived, may show this effect and could be useful models for screening transgene expression in founder lines. Consistent with the difference in transgene expression seen in undifferentiated neurospheres, differentiated cells derived from tauwt-expressing neurospheres expressed human tau in a higher proportion of cells than those derived from tauP301L-expressing neurospheres. Likely, non-transgene expressing progenitor cells gave rise to nontransgene expressing differentiated cells and transgene expressing progenitor cells differentiated into transgene expressing mature cells. Alternatively, tauP301L transgene expression may have decreased neural precursor survival. Since neurospheres proliferated and differentiated over several passages in both genotypes, and differentiated transgene expressing cell proportions mirrored that of the undifferentiated condition, evidence favors the former explanation. Some of the differences we saw between tauP301L and tauwt may stem from differences in transgene insertion sites. Neurosphere cultures from mouse lines expressing the same human tau variants, but at lower levels than rTg4510 and rTg21221, overlapped in the percentages of cells expressing human tau, but within the same range as our more extensively studies lines. It is likely that there are effects of transgene insertion site as well as the tau mutation. With the emergence of methodologies to culture neurospheres from human patients, experiments evaluating the culture system’s relevance and validity are crucial. Our data provide supporting evidence that neurospheres can reliably model phenotypes of their derived source over extended culture periods. The neurosphere culture system provides a robust assay for studying effects of external factors 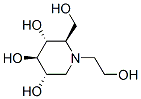 on the development and differentiation of the CNS, and the genetic susceptibility to neurological disorders. While it is unreasonable to expect that an in vitro system will fully recapitulate a complex disease process involving complex cell interactions, biologically relevant models that provide reproducible results are invaluable resources.
on the development and differentiation of the CNS, and the genetic susceptibility to neurological disorders. While it is unreasonable to expect that an in vitro system will fully recapitulate a complex disease process involving complex cell interactions, biologically relevant models that provide reproducible results are invaluable resources.
Phenotypes of neurosphere cultures isolated from individual fetuses at different times reflected
An engineered tau variant that mimics constitutively phosphorylated tau by replacing Ser/Thr residues with glutamate, did not develop tau aggregates, neuronal loss, or behavioral abnormalities. In contrast, mice expressing tauR406W, a variant that remains hypophosphorylated compared to tauwt even in aged mice, developed age-dependent tau aggregates and memory and behavioral abnormalities. The hypophosphorylated tauP301L species in neurospheres and young mice may represent free tau not associated with microtubules. Tau bound to MTs acquires more phosphorylation than free tau as demonstrated by in vitro phosphorylation of tau in the Folinic acid calcium salt pentahydrate presence or Pimozide absence of MTs. Many of the MAPT exon 10 missense mutations that cause dementia, including P301L, reduce the ability of tau to interact with MT, and tauwt displaces mutant tau from MTs. The absence of tauP301L aggregates or neurofibrillary tangles in neurospheres and in young mice, despite phosphorylation at many of the sites most frequently phosphorylated in AD and FTD, also may correspond to unbound tau as MT association has been implicated as an important step for tau nucleation. The quantity of NFTs correlates with disease severity. However, recent studies have dissociated NFTs from neuronal death and decreased memory function, and instead suggest a deleterious effect of soluble tau. Phosphorylation of mouse tau also reflects appropriate phosphorylation corresponding to the differentiation state. Heavily phosphorylated 3R mouse tau is know to bind to nucleolar organizing regions in dividing cells and was also observed in neurospheres, indicating that human tau did not inhibit either the cellular machinery or kinases involved. This reinforces the same conclusion coming from the recapitulation of the genetic differences in human tau phosphorylation seen in neurosphere culture. We observed differences in filopodia-spine densities between tauwt and tauP301L differentiated cells. Developmentally, dendritic spine morphology evolves from long thin filopodia-spines to mature spines of various morphologies; during this transformation, filopodia-spine density decreases. We observed a slightly higher filopodia-spine density in Map2 and TUJ-1 double-positive cells from tauP301L cultures than those derived from tauwt or cultures that did not express MAPT. While the level of transgene expression within individual cells did not affect filopodia-spine density, we have not ruled out an effect of the transgene insertion site. Interestingly, in a separate P301L mouse line that that harbors the mutation in the longest mouse tau isoform, 4R2N, driven by the Thy1 promoter, young mice with “hypophosphorylated” tau have enhanced learning and memory and increased Long Term Potentiation in the dentate gyrus 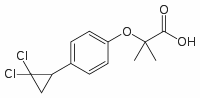 compared to controls. With age, the spine density of rTg4510 mice decreases and coincides with increased neuronal excitability. Whether or not the greater filopodiaspine density we observed in differentiated tauP301L cultures relates to enhanced LTP in young mice or neuronal vulnerability later in life in unknown, but warrants further exploration. We did not observe mislocalization of tauP301L to dendritic spines, as reported in aging mice or in transfected rat neuron cultures. We saw high levels of transgenic tau protein throughout neuritic projections in differentiated neurospheres from both human tau transgene genotypes. The presence of tau in the filopodia-spines of developing cells suggests that the phenomenon correlates with a dynamic morphology and does not necessarily indicate pathological injury.
compared to controls. With age, the spine density of rTg4510 mice decreases and coincides with increased neuronal excitability. Whether or not the greater filopodiaspine density we observed in differentiated tauP301L cultures relates to enhanced LTP in young mice or neuronal vulnerability later in life in unknown, but warrants further exploration. We did not observe mislocalization of tauP301L to dendritic spines, as reported in aging mice or in transfected rat neuron cultures. We saw high levels of transgenic tau protein throughout neuritic projections in differentiated neurospheres from both human tau transgene genotypes. The presence of tau in the filopodia-spines of developing cells suggests that the phenomenon correlates with a dynamic morphology and does not necessarily indicate pathological injury.